Включает в себя такие понятия, как поступление, движение воды в растениях и испарение ее.
Вода необходима растениям
Передвигается вода в растениях по клеткам коровой паренхимы до центрального цилиндра корня, затем по проводящей системе до листовой паренхимы и, наконец, по клеткам листовой паренхимы. На первом участке пути вода передвигается благодаря повышению сосущей силы клеток корня.
 Движение воды от корневого волоска в сторону центральных сосудов
Движение воды от корневого волоска в сторону центральных сосудов
Этот отрезок пути очень небольшой (доли миллиметра), но передвижение воды по этому участку очень затруднено, так как воде приходится преодолевать сопротивление слоев живой протоплазмы . Это сопротивление примерно равно 1 атм на 1 мм пути, поэтому передвижение воды по живым клеткам на более значительные расстояния не обеспечивало бы потребности растения в воде.
Действительно, растения , у которых не развита проводящая система , например мхи, (подробнее: ) имеют незначительные размеры и приспособлены к жизни только во влажных условиях. У наземных растений в процессе эволюции образовалась проводящая ткань, которая устанавливает сообщение между всасывающими воду корнями и испаряющими воду листьями.
Проводящая воду ткань
Проводящая воду ткань состоит из сосудов, или трахей , и трахеидов ; она начинается в центральном цилиндре корня, проходит через весь корень и стебель и заканчивается в виде тончайших разветвлений - жилок, пронизывающих всю листовую паренхиму.
Сосуды представляют собой мертвые трубки, образовавшиеся из живых клеток. В сосудах сохраняются поперечные перегородки на разном расстоянии (от нескольких миллиметров до метра в зависимости от вида растения) одна от другой.
Исчезновение перегородок даже на небольшом расстоянии в тысячи раз ускоряет передвижение воды. Трахеиды это длинные мертвые клетки с заостренными концами. При образовании сосудов и трахеид происходит утолщение и одревеснение их оболочек, вследствие чего они не сдавливаются под давлением окружающих их живых паренхимных клеток.
 Движение воды в древесных растениях
Движение воды в древесных растениях
Одревеснение, однако, никогда не бывает сплошным: на стенке сосудов остаются тонкие места - поры, по которым вода может перемещаться не только вверх по сосудам, но и в радиальном направлении.
Подъем воды по сосудам
Подъем воды по сосудам можно доказать следующим опытом. Если у срезанной и поставленной в воду ветки снять кольцо коры выше уровня воды, листья ее не завянут, так как сосуды расположены в древесине.
Движение воды по сосудам чаще всего направлено снизу вверх и называется поэтому восходящим током .

Последний отрезок пути водного тока по листовой паренхиме идет по живым
клеткам
. Вода передвигается осмотическим путем по клеткам мезофилла листа до последних клеток, граничащих с межклеточниками
. Этот отрезок пути, так же как и первый, очень короткий.
Если срезанную ветку растения герметически закрепить в стеклянной трубке, заполненной водой, и нижний конец ее опустить в сосуд со ртутью, то при испарении веткой воды ртуть в трубке будет подниматься.
Из этого опыта ясно, что передвижение воды по растению обусловлено главным образом транспирацией , (подробнее: ), а не только корневым давлением.
При испарении воды с поверхности листьев в клетках возникает сосущая сила. Величина ее тем больше, чем меньше воды остается в клетках листа. Эта возникающая сосущая сила поддерживает постоянное передвижение воды в растении.
Силы, приводящие воду в движение
Таким образом, силы, приводящие воду в движение , находятся по концам проводящей системы: нагнетающий воду корень, работа которого получила название нижнего концевого двигателя , и сила присасывания воды листьями - верхний концевой двигатель .
Оба двигателя действуют в одном направлении и могут заменять и дополнять друг друга. Во время сильной инсоляции летом и при, засухе водоснабжение растения идет за счет присасывающего действия транспирации.
Корневое давление
Когда же почва богата водой, а воздух водяными парами, подъем воды обеспечивается силой корневого давления, (подробнее: ). Следовательно, в зависимости от условий внешней среды главная роль принадлежит то одному, то другому концевому двигателю.
Водные нити не рвутся под влиянием своей тяжести, несмотря на то, что при сильной они находятся в состоянии натяжения. Это объясняется силой сцепления молекул воды, достигающей 300-350 атм, а так как в сосудах нет воздуха, то целостность водного тока не прерывается.
Скорость водного тока
Скорость водного тока зависит от строения проводящих воду элементов. Вода быстрее передвигается по сосудам, причем скорость движения ее зависит от диаметра сосудов: чем он меньше, тем медленнее будет передвигаться вода.
Движение воды в растениях происходит благодаря работе двух концевых двигателей, верхнего и нижнего, и сил сцепления, обеспечивающих целостность водных нитей.
В надземные части растения вода поднимается по ксилеме.
У хвойных она движется по трахеидам, у лиственных - по сосу-
дам и трахеидам. Эти клетки хорошо приспособлены для этой цели: они вытянуты в длину, лишены цитоплазмы и внутри полые, т.е. это как бы трубки для воды. Одревесневшие вторичные клеточные стенки достаточно прочны на разрыв, чтобы выдерживать огром-ную разность давлений, возникающую при подъеме воды к вер-шинам высоких деревьев. В ксилеме взрослых деревьев воду прово-дят в основном ее периферические слои - заболонь.
Движущей силой восходящего тока воды в проводящих эле-ментах ксилемы является градиент водного потенциала через рас-тение от почвы до атмосферы. Он поддерживается градиентом ос-мотического потенциала в клетках корня и транспирацией. Для поглощения воды корнями требуются затраты метаболической энергии. На транспирацию используется энергия солнечной ради-
ации. Транспирация - главная движущая сила восходящего тока воды, так как благодаря ей в ксилеме возникает отрицательное давление, т.е. натяжение. В силу сцепления (когезии) молекул воды друг с другом и действия сил прилипания (адгезии) ее к гидро-фильным стенкам сосудов столб воды в ксилеме непрерывен. Со-четание транспирации, когезии и натяжения обусловливает подъем воды в стволах высоких деревьев. У большинства древесных расте-ний водный ток в стволе передвигается по спирали. Это связано с макростроением древесного ствола. Линейная скоростьвосходящ-его тока колеблется от 1 - 6 м/ч у хвойных и рассеянно-сосуди-тых пород до 25 - 60 м/ч у кольцесосудистых. Он обеспечивает все живые клетки растения водой и минеральными элементами.
Содержание воды в древесине у большинства древесных растен-ий увеличивается от внутренней части ствола к внешней и от основания ствола к его вершине. В пределах кроны количество воды увеличивается от вершины к основанию. Резкие изменения овод-ненности древесины наблюдаются в течение года. Так, у хвойных древесных растений самая низкая влажность отмечается в летние месяцы, а самая высокая - зимой. Влажность ядровой древесины практически не меняется и остается самой низкой. У лиственных древесных пород отмечены два периода пониженной влажности- летний и во второй половине зимы и два повышенной - весен-ний во время сокодвижения и зимний - в первой половине зимы. В течение суток летом наиболее высокая влажность отмечается утром, а низкая - в полдень.
10.4. Транспирация
Основной орган транспирации - лист. В результате потери воды клетками листьев в них снижается водный потенциал, т.е. возрас-тает сосущая сила. Таким образом, верхний концевой двигатель, обеспечивающий передвижение воды вверх по растению, созда-ется и поддерживается высокой сосущей силой транспирирую-щих клеток листовой паренхимы. Физиологическая роль транспи-рации сводится к следующему: 1) повышает сосущую силу испа-ряющих клеток и создает непрерывный водный ток по растению;
2) способствует передвижению воды и растворенных в ней мине-ральных и частично органических веществ от корней к надзем-ным частям растения; 3) защищает листья от перегрева прямыми солнечными лучами; 4) препятствует полному насыщению кле-ток водой, так как при небольшом водном дефиците (до 5%) оптимизируется целый ряд процессов метаболизма.
Транспирация бывает устьичная, кутикулярная и корковая (пе-ридермальная). Испарение воды как физическое явление, т.е. пе-реход воды из жидкого состояния в парообразное, происходит в межклетниках листа с поверхности клеток мезофилла. Образую-щийся пар через устьица выделяется в атмосферу. Это устьичная транспирация.
Устьица - основные проводящие пути для водяного пара, СО, и О,. Они могут находиться на обеих сторонах листа, но есть виды, у которых устьица располагаются только на нижней стороне лис-та. В среднем число устьиц колеблется от 50 до 500 на 1 мм". Тран-спирация с поверхности листа через устьица идет почти с такой же скоростью, как с поверхности чистой воды.
Потери водяного пара через кутикулу листа при открытых ус-тьицах обычно очень незначительны по сравнению с общей транс-пирацией. Но если устьица закрыты, например во время засухи, кутикулярная транспирация приобретает важное значение в вод-ном режиме многих растений. Кутикулярная транспирация зави-
сит от толщины слоя кутикулы и сильно варьирует у разных видов.
У молодых листьев она составляет около половины от общей транс-пирации, у зрелых листьев, с более мощной кутикулой, - не превышает 10 %.
Некоторое количество воды выделяется в результате транспи-рации почек и репродуктивных органов. Иногда эти потери могут быть значительными: например, корзинки подсолнечника, коро-бочки мака и плоды перца транспирируют сильнее, чем листья данных растений в тех же условиях. Вода испаряется с поверхнос-ти ветвей и стволов древесных растений через чечевички и окру-жающие их слои пробки. Это пробковая, или перидермальная, транс-пираиия. Вследствие транспирации ветвей и почек в зимнее время нередко наблюдаются случаи, когда значительные потери воды приводят к суховершинности древесных растений.
Скорость транспирации и газообмен в целом регулируются ус-тьицами. Степень раскрытости устьиц зависит от освещенности, оводненности тканей листа, концентрации СО2 в межклетниках и " .других факторов. В зависимости от факторов, запускающих двига-тельный механизм (свет или начинающийся водный дефицит в тканях листа), различают фото- и гидроактивное движение усть-иц. На свету в хлоропластах замыкающих клеток начинается фото-синтез, что вызывает уменьшение содержания СО2, накопивше-гося за ночь в клетках. При этом происходит накопление АТФ и, превращение крахмала в сахар,за счет которого активизируются
ионные насосы, перекачивающие калий из соседних клеток. Бла-годаря этому резко повышается сосущая сила устьичных клеток, всасывающих воду и увеличивающих тургор. Все это способствует открыванию устьиц. При возникновении водного дефицита повы-шается содержание одного из гормонов - абсцизовой кислоты; , под ее действием происходит отток и других растворенных ве-ществ, что приводит к закрыванию устьиц. Этот механизм позво-ляет защитить растение от излишней потери воды.
Показателем транспирации является ее интенсивность - коли-честно воды, испаренной в единицу времени на единицу сырой или сухой массы или поверхности листа (мг/дм2ч, г/м2ч или мг/г ч).
Количество граммов сухой массы, образовавшейся при испаре-нии 1 л воды, называется продуктивностью транспирации. Под эвапотранспирацией понимается сумма транспирационных потерь всеми растениями сообщества плюс физическое испарение (эва-порация) с поверхности почвы и растений, в частности со ство-лов и ветвей деревьев. Для лесных площадей центральных районов европейской части России транспирация древостоя в среднем соста-вляет 50 - 60 % эвапотранспирации, напочвенного покрова - 15- " 25%, испарение с поверхности почвы и растений - 25 - 35%.
Транспирацию кроны дерева используют для подсушки древе-сины после рубки. Свежесрубленная древесина ряда древесных пород (лиственница, береза, осина и др.) настолько тяжела, что тонет при сплаве. Вместе с тем более сухая и, следовательно, бо-лее легкая древесина тех же пород с успехом сплавляется на дале-кие расстояния. Для подсушки срубленное дерево оставляют ле-жать в лесу с кроной в течение 10 - 15 суток. Дерево продолжает жить за счет внутренних запасов воды и питательных веществ, а листья транспирировать. Количество свободной воды в стволе по-нижается. Уменьшение массы 1 мз древесины за указанное время составляет 25 - 30%, что резко повышает ее плавучесть. Облегча-ются также ее трелевка и транспортировка. Известно, что после сплава предварительно подсушенная древесина быстрее сохнет, чем не подсушенная до сплава.
На интенсивность транспирации влияет ряд факторов: доступ-ность воды корням растений, влажность воздуха, температура, ветер. При недостатке воды в почве интенсивность транспирации древесных растений заметно снижается. На затопленной почве этот процесс, несмотря на обилие воды, также снижен у деревьев при-мерно в 1,5 - 2 раза, что связано с плохой аэрацией корневых систем. Уменьшается транспирация и при сильном охлаждении почвы в связи со снижением скорости поглощения воды. Недоста-ток или избыток воды, засоление или холодная почва действуют на интенсивность транспирации не сами по себе, а через их вли-яние на поглощение воды корневыми системами.
Свет и влажность воздуха сильно воздействуют на транспира-цию. Свет увеличивает открытость устьиц. Интенсивность транс-пирации даже на рассеянном свету повышается на 30 - 40%, В темноте растения транспирируют в десятки раз слабее, чем при полном солнечном освещении. Повышение относительной влаж-ности приводит к резкому снижению интенсивности транспира-ции всех пород. Согласно закону Дальтона, количество испаряе-мой воды прямо пропорционально дефициту насыщения воздуха водяными парами.
Температура воздуха действует на транспирацию прямо и кос-венно. Прямое воздействие связано с нагреванием листа, а кос-венное - через изменение упругости паров, насыщающих про-странство. При повышении температуры количество пара в возду-хе уменьшается и транспирация усиливается. Ветер способствует повышению транспирации благодаря уносу паров воды от листь-ев, создавая недонасыщение воздуха у их поверхности.
В природе всегда действует комплекс факторов. В ходе дня ме-няются освещенность, температура и влажность воздуха, что ве-дет к изменению интенсивности транспирации (рис. 10.2). При умеренных температуре и влажности содержание воды в листьях уменьшается незначительно - на 10 - 15%. В жаркий день овод-ненность листьев снижается по сравнению с нормой до 25% и более.

Рис. 10.2. Дневной ход транспира-ции при различной влагообеспе-ченности растений:
А - испарение со свободной водной по-верхности; Б - транспирация при дос-таточной влагообеспеченности; В - при недостатке влаги в полдень; Г - при глубоком водном дефиците; Д - во вре-мя длительной засухи.
Различают дневной и остаточный водный дефицит. Дневной водный дефицит наблюдается в полуденные часы летнего дня. Как правило, он существенно не нарушает жизнедеятельность расте-ний. Остаточный водный дефицит наблюдается на рассвете и сви-детельствует о том, что водные запасы листа за ночь восстанови-лись лишь частично вследствие низкой влажности почвы. При этом растения сначала сильно завядают, а затем при длительной засухе могут погибнуть.
Контрольные вопросы:
1. Из чего складывается водный режим растения?
2. Как происходит поглощение воды корнями?
3. В чем проявляется корневое давление?
4. Какие формы почвенной влаги доступны для растения?
5. Как осуществляется подъем воды к вершинам высоких деревьев?
6. Что такое транспирация и какая она бывает?
7. Как растение регулирует транспирацию?
8. Какие факторы внешней среды оказывают влияние на интенсив-ность транспирации?
МИНЕРАЛЬНОЕ ПИТАНИЕ.
Филогенетически корень возник позже стебля и листа - в связи с переходом растений к жизни на суше и вероятно, произошёл от корнеподобных подземных веточек. У корня нет ни листьев, ни в определённом порядке расположенных почек. Для него характерен верхушечный рост в длину, боковые разветвления его возникают из внутренних тканей, точка роста покрыта корневым чехликом. Корневая система формируется на протяжении всей жизни растительного организма. Иногда корень может служить местом отложения в запас питательных веществ. В таком случае он видоизменяется.
Виды корней
Главный корень образуется из зародышевого корешка при прорастании семени. От него отходят боковые корни.
Придаточные корни развиваются на стеблях и листьях.
Боковые корни представляют собой ответвления любых корней.
Каждый корень (главный, боковые, придаточные) обладает способностью к ветвлению, что значительно увеличивает поверхность корневой системы, а это способствует лучшему укреплению растения в почве и улучшению его питания.
Типы корневых систем
Различают два основных типа корневых систем: стержневая, имеющая хорошо развитый главный корень, и мочковатая. Мочковатая корневая система состоит из большого числа придаточных корней, одинаковых по величине. Вся масса корней состоит из боковых или придаточных корешков и имеет вид мочки.
Сильно разветвлённая корневая система образует огромную поглощающую поверхность. Например,
- общая длина корней озимой ржи достигает 600 км;
- длина корневых волосков — 10 000 км;
- общая поверхность корней — 200 м 2 .
Это во много раз превышает площадь надземной массы.
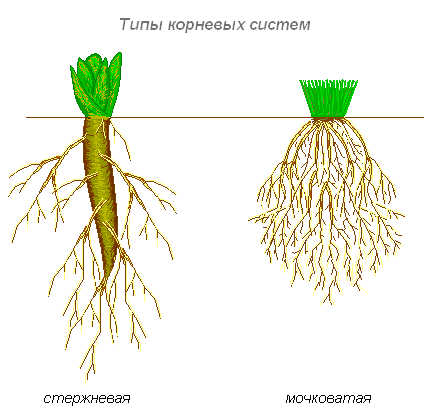
Если у растения хорошо выражен главный корень и развиваются придаточные корни, то формируется корневая система смешанного типа (капуста, помидор).
Внешнее строение корня. Внутреннее строение корня
Зоны корня
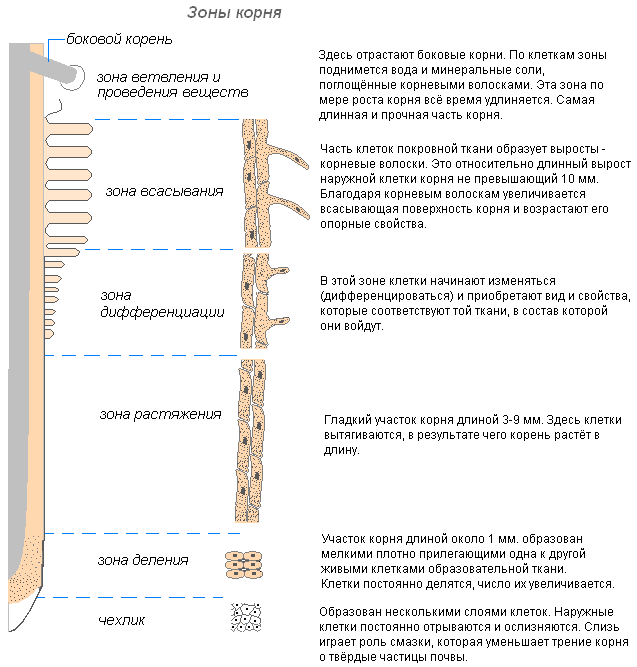
Корневой чехлик
Корень растёт в длину своей верхушкой, где находятся молодые клетки образовательной ткани. Растущая часть покрыта корневым чехликом, защищающим кончик корня от повреждений, и облегчает продвижение корня в почве во время роста. Последняя функция осуществляется благодаря свойству внешних стенок корневого чехлика покрываться слизью, что уменьшает трение между корнем и частичками почвы. Могут даже раздвигать частички почвы. Клетки корневого чехлика живые, часто содержат зёрна крахмала. Клетки чехлика постоянно обновляются за счёт деления. Участвует в положительных геотропических реакциях (направление роста корня к центру Земли).

Клетки зоны деления активно делятся, протяженность этой зоны у разных видов и у разных корней одного и того же растения неодинакова.
За зоной деления расположена зона растяжения (зона роста). Протяжённость этой зоны не превышает нескольких миллиметров.
По мере завершения линейного роста наступает третий этап формирования корня — его дифференциация, образуется зона дифференциации и специализации клеток (или зона корневых волосков и всасывания). В этой зоне уже различают наружный слой эпиблемы (ризодермы) с корневыми волосками, слой первичной коры и центральный цилиндр.
Строение корневого волоска
Корневые волоски — это сильно удлинённые выросты наружных клеток, покрывающих корень. Количество корневых волосков очень велико (на 1 мм 2 от 200 до 300 волосков). Их длина достигает 10 мм. Формируются волоски очень быстро (у молодых сеянцев яблони за 30-40 часов). Корневые волоски недолговечны. Они отмирают через 10-20 дней, а на молодой части корня отрастают новые. Это обеспечивает освоение корнем новых почвенных горизонтов. Корень непрерывно растёт, образуя всё новые и новые участки корневых волосков. Волоски могут не только поглощать готовые растворы веществ, но и способствовать растворению некоторых веществ почвы, а затем всасывать их. Участок корня, где корневые волоски отмерли, некоторое время способен всасывать воду, но затем покрывается пробкой и теряет эту способность.

Оболочка волоска очень тонкая, что облегчает поглощение питательных веществ. Почти всю клетку волоска занимает вакуоль, окружённая тонким слоем цитоплазмы. Ядро находится в верхней части клетки. Вокруг клетки образуется слизистый чехол, который содействует склеиванию корневых волосков с частицами почвы, что улучшает их контакт и повышает гидрофильность системы. Поглощению способствует выделение корневыми волосками кислот (угольной, яблочной, лимонной), которые растворяют минеральные соли.
Корневые волоски играют и механическую роль — они служат опорой верхушке корня, которая проходит между частичками почвы.
Под микроскопом на поперечном срезе корня в зоне всасывания видно его строение на клеточном и тканевом уровнях. На поверхности корня — ризодерма, под ней — кора. Наружный слой коры — экзодерма, вовнутрь от неё — основная паренхима. Её тонкостенные живые клетки выполняют запасающую функцию, проводят растворы питательных веществ в радиальном направлении — от всасывающей ткани к сосудам древесины. В них же происходит синтез ряда жизненно важных для растения органических веществ. Внутренний слой коры — эндодерма. Растворы питательных веществ, поступающие из коры в центральный цилиндр через клетки эндодермы, проходят только через протопласт клеток.

Кора окружает центральный цилиндр корня. Она граничит со слоем клеток, долго сохраняющих способность к делению. Это перицикл. Клетки перицикла дают начало боковым корням, придаточным почкам и вторичным образовательным тканям. Вовнутрь от перицикла, в центре корня, находятся проводящие ткани: луб и древесина. Вместе они образуют радиальный проводящий пучок.
Проводящая система корня проводит воду и минеральные вещества из корня в стебель (восходящий ток) и органические вещества из стебля в корень (нисходящий ток). Состоит она из сосудисто-волокнистых пучков. Основными слагаемыми частями пучка являются участки флоэмы (по ним вещества передвигаются к корню) и ксилемы (по которым вещества передвигаются от корня). Основные проводящие элементы флоэмы — ситовидные трубки, ксилемы — трахеи (сосуды) и трахеиды.
Процессы жизнедеятельности корня
Транспорт воды в корне
Всасывание воды корневыми волосками из почвенного питательного раствора и проведение её в радиальном направлении по клеткам первичной коры через пропускные клетки в эндодерме к ксилеме радиального проводящего пучка. Интенсивность поглощения воды корневыми волосками называется сосущей силой (S), она равна разнице между осмотическим (P) и тургорным (T) давлением: S=P-T.
Когда осмотическое давление равно тургорному (P=T), то S=0, вода перестаёт поступать в клетку корневого волоска. Если концентрация веществ почвенного питательного раствора будет выше, чем внутри клетки, то вода будет выходить из клеток и наступит плазмолиз — растения завянут. Такое явление наблюдается в условиях сухости почвы, а также при неумеренном внесении минеральных удобрений. Внутри клеток корня сосущая сила корня возрастает от ризодермы по направлению к центральному цилиндру, поэтому вода движется по градиенту концентрации (т. е. из места с большей её концентрацией в место с меньшей концентрацией) и создаёт корневое давление, которое поднимает столбик воды по сосудам ксилемы, образуя восходящий ток. Это можно обнаружить на весенних безлистных стволах, когда собирают «сок», или на срезанных пнях. Истекание воды из древесины, свежих пней, листьев, называется «плачем» растений. Когда распускаются листья, то они тоже создают сосущую силу и притягивают воду к себе — образуется непрерывный столбик воды в каждом сосуде — капиллярное натяжение. Корневое давление является нижним двигателем водного тока, а сосущая сила листьев — верхним. Подтвердить это можно с помощью несложных опытов.
Всасывание воды корнями
Цель: выяснить основную функцию корня.
Что делаем: растение, выращенное на влажных опилках, отряхнём его корневую систему и опустим в стакан с водой его корни. Поверх воды для защиты её от испарения нальём тонкий слой растительного масла и отметим уровень.

Что наблюдаем: через день-два вода в ёмкости опустилась ниже отметки.

Результат: следовательно, корни всосали воду и подали её наверх к листьям.
Можно ещё проделать один опыт, доказывающий всасывание питательных веществ корнем.
Что делаем: срежем у растения стебель оставив пенёк высотой 2-3 см. На пенёк наденем резиновую трубку длиной 3 см, а на верхний конец наденем изогнутую стеклянную трубку высотой 20-25 см.
Что наблюдаем: вода в стеклянной трубке поднимается, и вытекает наружу.
Результат: это доказывает, что воду из почвы корень всасывает в стебель.

А влияет ли температура воды на интенсивность всасывания корнем воды?
Цель: выяснить, как температура влияет на работу корня.
Что делаем: один стакан должен быть с тёплой водой (+17-18ºС), а другой с холодной (+1-2ºС).
Что наблюдаем: в первом случае вода выделяется обильно, во втором — мало, или совсем приостанавливается.
Результат: это является доказательством того, что температура сильно влияет на работу корня.

Тёплая вода активно поглощается корнями. Корневое давление повышается.

Холодная вода плохо поглощается корнями. В этом случае корневое давление падает.
Минеральное питание
Физиологическая роль минеральных веществ очень велика. Они являются основой для синтеза органических соединений, а также факторами, которые изменяют физическое состояние коллоидов, т.е. непосредственно влияют на обмен веществ и строение протопласта; выполняют функцию катализаторов биохимических реакций; воздействуют на тургор клетки и проницаемость протоплазмы; являются центрами электрических и радиоактивных явлений в растительных организмах.
Установлено, что нормальное развитие растений возможно только при наличии в питательном растворе трёх неметаллов — азота, фосфора и серы и — и четырёх металлов — калия, магния, кальция и железа. Каждый из этих элементов имеет индивидуальное значение и не может быть заменён другим. Это макроэлементы, их концентрация в растении составляет 10 -2 –10%. Для нормального развития растений нужны микроэлементы, концентрация которых в клетке составляет 10 -5 –10 -3 %. Это бор, кобальт, медь, цинк, марганец, молибден др. Все эти элементы есть в почве, но иногда в недостаточном количестве. Поэтому в почву вносят минеральные и органические удобрения.
Растение нормально растёт и развивается в том случае, если в окружающей корни среде будут содержаться все необходимые питательные вещества. Такой средой для большинства растений является почва.
Дыхание корней
Для нормального роста и развития растения необходимо чтобы к корню поступал свежий воздух. Проверим, так ли это?
Цель: нужен ли воздух корню?
Что делаем: возьмём два одинаковых сосуда с водой. В каждый сосуд поместим развивающие проростки. Воду в одном из сосудов каждый день насыщаем воздухом с помощью пульверизатора. На поверхность воды во втором сосуде нальём тонкий слой растительного масла, так как оно задерживает поступление воздуха в воду.

Что наблюдаем: через некоторое время растение во втором сосуде перестанет расти, зачахнет, и в конце концов погибнет.

Результат: гибель растения наступает из-за недостатка воздуха, необходимого для дыхания корня.
Видоизменения корней
У некоторых растений в корнях откладываются запасные питательные вещества. В них накапливаются углеводы, минеральные соли, витамины и другие вещества. Такие корни сильно разрастаются в толщину и приобретают необычный внешний вид. В формировании корнеплодов участвуют и корень, и стебель.
Корнеплоды
Если запасные вещества накапливаются в главном корне и в основании стебля главного побега, образуются корнеплоды (морковь). Растения, образующие корнеплоды, в основном двулетники. В первый год жизни они не цветут и накапливают в корнеплодах много питательных веществ. На второй — они быстро зацветают, используя накопленные питательные вещества и образуют плоды и семена.
Корневые клубни

У георгина запасные вещества накапливаются в придаточных корнях, образуя корневые клубни.
Бактериальные клубеньки
Своеобразно изменены боковые корни у клевера, люпина, люцерны. В молодых боковых корешках поселяются бактерии, что способствует усвоению газообразного азота почвенного воздуха. Такие корни приобретают вид клубеньков. Благодаря этим бактериям эти растения способны жить на бедных азотом почвах и делать их более плодородными.
Ходульные

У пандуса, произрастающего в приливно-отливной зоне, развиваются ходульные корни. Они высоко над водой удерживают на зыбком илистом грунте крупные облиственные побеги.
Воздушные

У тропических растений, живущих на ветвях деревьев, развиваются воздушные корни. Они часто встречаются у орхидей, бромелиевых, у некоторых папоротников. Воздушные корни свободно висят в воздухе, не достигая земли и поглощая попадающую на них влагу от дождя или росы.
Втягивающие

У луковичных и клубнелуковичных растений, например у крокусов, среди многочисленных нитевидных корней имеется несколько более толстых, так называемых втягивающих, корней. Сокращаясь, такие корни втягивают клубнелуковицу глубже в почву.
Столбовидные

У фикуса развиваются столбовидные надземные корни, или корни-подпорки.
Почва как среда обитания корней
Почва для растений является средой, из которой оно получает воду и элементы питания. Количество минеральных веществ в почве зависит от специфических особенностей материнской горной породы, деятельности организмов, от жизнедеятельности самих растений, от типа почвы.
Почвенные частицы конкурируют с корнями за влагу, удерживая её своей поверхностью. Это так называемая связанная вода, которая подразделяется на гигроскопическую и плёночную. Удерживается она силами молекулярного притяжения. Доступная растению влага представлена капиллярной водой, которая сосредоточена в мелких порах почвы.
Между влагой и воздушной фазой почвы складываются антагонистические отношения. Чем больше в почве крупных пор, тем лучше газовый режим этих почв, тем меньше влаги удерживает почва. Наиболее благоприятный водно-воздушный режим поддерживается в структурных почвах, где вода и воздух находятся одновременно и не мешают друг другу — вода заполняет капилляры внутри структурных агрегатов, а воздух — крупные поры между ними.
Характер взаимодействия растения и почвы в значительной степени связан с поглотительной способностью почвы — способностью удерживать или связывать химические соединения.
Микрофлора почвы разлагает органические вещества до более простых соединений, участвует в формировании структуры почвы. Характер этих процессов зависит от типа почвы, химического состава растительных остатков, физиологических свойств микроорганизмов и других факторов. В формировании структуры почвы принимают участие почвенные животные: кольчатые черви, личинки насекомых и др.
В результате совокупности биологических и химических процессов в почве образуется сложный комплекс органических веществ, который объединяют термином «гумус».
Метод водных культур
В каких солях нуждается растение, и какое влияние оказывают они на рост и развитие его, было установлено на опыте с водными культурами. Метод водных культур — это выращивание растений не в почве, а в водном растворе минеральных солей. В зависимости от поставленной цели в опыте можно исключить отдельную соль из раствора, уменьшить или увеличить ее содержание. Было выяснено, что удобрения, содержащие азот, способствуют росту растений, содержащие фосфор — скорейшему созреванию плодов, а содержащие калий — быстрейшему оттоку органических веществ от листьев к корням. В связи с этим содержащие азот удобрения рекомендуется вносить перед посевом или в первой половине лета, содержащие фосфор и калий — во второй половине лета.
С помощью метода водных культур удалось установить не только потребность растения в макроэлементах, но и выяснить роль различных микроэлементов.
В настоящее время известны случаи, когда выращивают растения методами гидропоники и аэропоники.
Гидропоника — выращивание растений в сосудах, заполненных гравием. Питательный раствор, содержащий необходимые элементы, подаётся в сосуды снизу.
Аэропоника — это воздушная культура растений. При этом способе корневая система находится в воздухе и автоматически (несколько раз в течение часа) опрыскивается слабым раствором питательных солей.
Вода, поступившая в клетки корня под влиянием разности водных потенциалов, которые возникают благодаря транспирации и корневого давления, передвигается до проводящих элементов ксилемы. Согласно современным представлениям, вода в корневой системе может перемещаться в радиальном направлении тремя путями:
- апопластическим,
- симпластическим,
- трансмембранным.
При транспорте по апопласту вода передвигается по клеточным стенкам, не проходя через мембраны. При симпластном т ранспорте вода проникает в клетку через полупроницаемую мембрану и далее перемещается по протопластам клеток, которые соединены между собой многочисленными плазмодесмами. При трансмембранном транспорте вода перетекает через клетки и при этом проходит, по крайней мере, две плазматические мембраны. В последнее время много внимания уделяется аквапоринам - мембранным белкам, образующим в мембранах специализированные водные каналы и определяющим проницаемость для воды. Эксперименты показали, что передвижение воды по коре корня идет главным образом по апопласту, где она встречает меньшее сопротивление, и лишь частично по симпласту (С. Френч). Апопластный путь прерывается в эндодерме в связи с наличием поясков Каспари . Вместе с тем в апикальной части суберинизация отсутствует, поэтому вода легко проникает через эндодерму. Кроме того, в суберинизированных частях корня вода может проходить через пропускные клетки.
Для транспорта в сосуды ксилемы, вода должна пройти через полупроницаемую мембрану клеток эндодермы. Таким образом, мы имеем дело как бы с осмометром, у которого полупроницаемая мембрана расположена в клетках эндодермы. Вода устремляется через эту мембрану. В сторону меньшего (более отрицательного) водного потенциала. Далее вода поступает в сосуды ксилемы.
По вопросу о причинах, вызывающих секрецию воды в сосуды ксилемы, имеются различные суждения. Согласно гипотезе Крафтса , это следствие выброса солей в сосуды ксилемы, в результате чего там создается повышенная их концентрация, и водный потенциал становится более отрицательным . Предполагается, что в результате активного поступления соли накапливаются в клетках корня. Однако интенсивность дыхания в клетках, окружающих сосуды ксилемы (перицикл), очень низкая, и они не удерживают соли, которые благодаря этому десорбируются в сосуды. Транспорт воды в корне зависит от интенсивности процесса дыхания . При помещении растений в условия, тормозящие дыхание корней (низкая температура, анаэробиоз или наличие дыхательных ядов), они транспортируют меньше воды.
Дальнейшее передвижение воды идет по сосудистой системе корня, стебля и листа. Проводящие элементы ксилемы состоят из сосудов и трахеид. Опыты с кольцеванием показали, что восходящий ток воды по растению движется в основном по ксилеме. В проводящих элементах ксилемы вода встречает незначительное сопротивление, что, естественно, облегчает передвижение воды на большие расстояния. Правда, некоторое количество воды передвигается и вне сосудистой системы. Вне ксилемы движется всего от 1 до 10% общего потока воды. Из сосудов стебля вода попадает в сосуды листа. Вода движется из стебля через черешок или листовое влагалище в лист. В листовой пластинке водопроводящие сосуды расположены в жилках. Жилки, постепенно разветвляясь, становятся все более мелкими. Чем гуще сеть жилок, тем меньшее сопротивление встречает вода при передвижении к клеткам мезофилла листа. Именно поэтому густота жилкования листа считается одним из важнейших признаков ксероморфной структуры - отличительной чертой растений, устойчивых к засухе.
Вся вода в клетке находится в равновесном состоянии . Иначе говоря, в смысле насыщенности водой имеется равновесие между вакуолью, цитоплазмой и клеточной оболочкой, их водные потенциалы равны. В связи с этим, как только в силу процесса транспирации возникает ненасыщенность водой клеточных стенок паренхимных клеток, она сейчас же передается внутрь клетки, водный потенциал которой падает. Вода передвигается от клетки к клетке благодаря градиенту водного потенциала . По-видимому, передвижение воды от клетки к клетке в листовой паренхиме идет не по симпласту, а в основном по клеточным стенкам , где сопротивление значительно меньше. По сосудам вода движется благодаря создающемуся в силу транспирации градиенту водного потенциала, градиенту свободной энергии (от системы с большей свободой энергии к системе с меньшей). Распределение водных потенциалов, которое и вызывает передвижение воды: водный потенциал почвы (-0,5 бар), корня (-2 бар), стебля (-5 бар), листьев (-15 бар), воздуха при относительной влажности 50% (-1000 бар).
Восходящий ток воды может осуществляться и при отсутствии транспирации. К этому же выводу приводят опыты, показывающие ритмическое секретирование устьичными клетками жидкой воды, а также зависимость передвижения воды от эндогенной энергии, поставляемых процессом дыхания. Это позволяет считать, что движущая сила транспорта воды в растении является суммой двух весьма различных по своей природе составляющих, условно названных метаболической и осмотической . Осмотическая составляющая представлена в корнях сугубо осмотическими явлениями, в стебле и листьях - гидростатической тягой, создаваемой градиентом водного потенциала в системе почва - растение - атмосфера. Температурный коэффициент Q этой составляющей близок к единице, что характерно для простых физических процессов . В то же время Q процесса транспорта воды значительно выше. Столь высокий температурный коэффициент свойственен сложным процессам цепного характера. Очевидно, транспорт воды за счет метаболической составляющей относится к разряду именно таких процессов. Согласно имеющимся экспериментальным данным, в формировании метаболической составляющей непосредственное участие принимают контрактильные системы паренхимных клеток (т. е. белки цитоскелета). Они могут играть роль сфинктеров, регулирующих просветы водных каналов (открывание-закрывание) в плазмодесмах. Благодаря их работе индуцируются ритмические микроколебания внутриклеточного (внутрисимпластного) гидростатического давления, которые внешне выражаются в короткопериодных (1-3 мин) автоколебаниях поступления воды в клетку (симпласт, ткань, орган) и водоотдачи (т. е. микропульсациях). Итак, процесс транспорта воды состоит из двух последовательных, ритмично чередующихся, относительно самостоятельных, хотя и тесно взаимосвязанных фаз:
- фазы сокращения, во время которой вода выделяется (выталкивается, секретируется),
- последующей фазы расслабления, во время которой вода поглощается.
Выделение воды вызывает падение тургора и водного потенциала в целом, создавая предпосылку для поглощения следующей порции воды, вновь приводящего к возрастанию водного потенциала вплоть до того, что он из отрицательного становится положительным. После этого происходит новое сокращение. Именно фаза сокращения происходит с участием контрактильных систем и требует затраты энергии. Таким образом, вода поглощается и выделяется по градиенту водного потенциала , транспорт воды в термодинамическом понимании является пассивным .
Возникающие за счет ритмической деятельности внутриклеточного сократительного аппарата микроколебания гидростатического давления паренхимных клеток являются механизмом, создающим локальные градиенты водного потенциала на пути водного тока и тем самым регулирующим скорость этого тока. Именно благодаря этому формируется метаболическая составляющая движущей силы транспорта воды в растении, играющая решающую роль в общей системе эндогенной регуляции. Под влиянием ингибиторов контрактильных систем или окислительного фосфорилирования (т. е. при нарушении энергоснабжения) противофазность исчезает, автоколебания затухают и транспорт воды тормозится (В.Н. Жолкевич).
Вся вода в растении представляет единую взаимосвязанную систему. Поскольку между молекулами воды имеются силы сцепления (когезия), вода поднимается на высоту, значительно большую 10 м. Расчеты показали, что благодаря наличию сродства между молекулами воды силы сцепления достигают величины -30 бар. Это такая сила, которая позволяет поднять воду на высоту, равную 120 м, без разрыва водных нитей, что примерно составляет максимальную высоту деревьев. Силы сцепления существуют и между водой и стенками сосудов - адгезия . Стенки проводящих элементов ксилемы эластичны. Даже при недостатке воды связь между молекулами воды и стенками сосудов не нарушается. Это подтверждается исследованиями по изменению толщины стебля травянистых растений. Определения показали, что в полуденные часы толщина стебля растений уменьшается. Если перерезать стебель, то сосуды сразу расширяются и воздух поступает в них.
Степень натяжения водных нитей в сосудах зависит от соотношения процессов поглощения и испарения воды. Все это позволяет растительному организму поддерживать единую водную систему и не обязательно восполнять каждую каплю испаряемой воды. Таким образом, при нормальном водоснабжении создается непрерывность воды в системе почва - растение - атмосфера. В том случае, если в отдельные членики сосудов попадает воздух (эмболия), они, по-видимому, выключаются из общего тока проведения воды. Таков путь воды по растению и его основные движущие силы.
Вода движется в растении по градиенту водного потенциала. Вода, поглощенная корневыми волосками и другими клетками эпидермиса, из клеток внешней части корня перемещается к ксилеме, занимающей центральную часть корня (рис. 6.8). Главным путем диффузии воды во внешней части корня служит апопласт — непрерывная совокупность клеточных стенок. Однако в эндодерме (цилиндрическом слое клеток, окружающем проводящую ткань) свободная диффузия по клеточным стенкам наталкивается на преграду — водонепроницаемый пробковый слой пояска Каспари. Вода должна изменить здесь свой путь и пройти сквозь мембрану и протопласт клеток эндодермы, играющей, таким образом, роль осмотического барьера между корой корня и его центральным цилиндром. У однодольных пробковеют также и внутренние тангенциальные стенки клеток, но эти стенки пронизаны порами, по которым, как по каналам, может проходить вода.
Рис. 6.7. Корешок проростка горчицы. Многочисленные тонкие выросты — корневые волоски
По ксилеме вода поднимается в надземные части растения. Ксилема состоит из нескольких типов клеток. Вода движется в ней главным образом по сосудам и трахеидам (рис. 2.6 и 6.9). И те и другие клетки прекрасно приспособлены для этой цели: они вытянуты в длину, лишены живого содержимого и внутри полые, т. е. это как бы трубки для воды. Одревесневшие вторичные клеточные стенки достаточно прочны на разрыв, чтобы выдерживать огромную разность давлений, возникающую при подъеме воды к вершинам высоких деревьев. Торцевые, а иногда и боковые стенки члеников сосудов перфорированы; сосуды, состоящие из соединенных конец в конец члеников, образуют длинные трубки, по которым легко проходит вода с растворенными в ней минеральными веществами. В трахеидах нет перфораций, и вода, для того чтобы попасть из одной трахеиды в другую, должна пройти через их торцевые стенки; однако трахеиды — очень длинные клетки, а потому и эта конструкция достаточно хорошо приспособлена для проведения воды. У цветковых растений есть и сосуды, и трахеиды; у более при-митивных форм сосудов, как правило, нет.

Рис. 6.8. Поступление воды из почвы в корень. Вода может перемещаться как по апопласту, так и по симпласту до тех пор, пока она не достигнет эндодермы. Дальнейшее передвижение по апопласту оказывается невозможным; здесь этот путь перекрыт барьером — пояском Каспари. В нижней части рисунка показан при большом увеличении поясок Каспари — водонепроницаемый барьер, заставляющий воду покинуть апопласт и устремиться через мембраны клеток эндодермы в симпласт
Листовые жилки, состоящие из тяжей ксилемы и флоэмы, образуют в листе настолько густую сеть, что любая его клетка оказывается достаточно близко от источника воды (рис. 3.9). Из ксилемы вода диффундирует в стенки клеток мезофилла. Таким образом, вода в жидкой фазе заполняет весь путь от почвы — через корень и стебель — до клеток мезофилла в листе. Суммарный поток воды направлен всегда в сторону меньшего водного потенциала, т. е. ψ максимален в почве, несколько ниже в клетках корня и самый низкий в клетках, примыкающих к эпидермимису листа. Малая величина ψ в этих последних клетках объясняется главным образом испарением воды с поверхности листа, т. е. транспирацией (которой посвящен следующий раздел). Табл. 6.2 дает представление о градиентах, обусловливающих движение воды в растении от ее поступления из почвы до испарения в атмосферу.

Рис. 6.9. А. Поперечный срез листового черешка клещевины (Ricinus communis) . Видны проводящие элементы ксилемы со спиральными утолщениями. Благодаря такому строению трубка может растягиваться по мере того, как черешок растет. (Диаметр более крупной трубки равен приблизительно 15 мкм.) Б. Продольный срез стебля Ricinus, на котором виден сосуд ксилемы и перфорированные торцевые стенки двух члеников сосуда в месте их соединения. Видны эллиптические и округлые окаймленные поры с замыкающей пленкой, но без торуса такие поры типичны для двудольных. (Диаметр сосуда равен приблизительно 21 мкм.) (С любезного разрешения Milburn J. A., Glasgow University; фотографии сделаны при помощи сканирующего электронного микроскопа.)

Таблица 6.2 Примерные значения водного потенциала (ψ) и разности водных потенциалов (Δψ) для гипотетической системы почва — растение — воздух. [Предполагаемые условия — сравнительно небольшое дерево, хорошо увлажненная почва, относительная влажность воздуха около 50% при 22 °С (ψ=-1000 бар)1)]
Вода переходит из растения в окружающий воздух главным образом в парообразном состоянии. В мезофилле листа имеются обширные межклеточные пространства, и каждая клетка мезофилла хотя бы одной своей стороной граничит с таким межклетником. Вследствие испарения воды с влажных клеточных стенок воздух в межклетниках насыщен водяными парами, и часть этих паров теряется — выходит наружу. Поскольку у большинства растений клетки эпидермиса покрыты воскообразной водонепроницаемой кутикулой, водяные пары выходят из листа в атмосферу главным образом через устьица (рис. 6.10).

Рис. 6.10. Поперечный срез листа, на котором видно открытое устьице (У) с подустьичной воздушной полостью (ВП). Обратите внимание на то, что крупные, заполненные воздухом межклетники пронизывают всю толщу листа
Движение воды в растениях
Основным источником влаги является вода, находящаяся в почве, и основным органом поглощения воды в растение - корневая система. Роль этого органа, прежде всего, заключается в том, что благодаря огромной поверхности обеспечивается поступление воды в растение из большего объема почвы.
Сформировавшаяся корневая система представляет собой сложный орган с хорошо дифференцированной внешней и внутренней структурой. Корневая система имеет поглощающую или всасывающую зону — это зона корневых волосков. Поступив в клетку корневого волоска, вода становится частью живой системы — клетки растения — и подчиняется закономерностям, действующим в живой клетке.
Передвижение по растению определяется двумя основными двигателями водного потока в растении: нижним двигателем водного потока или корневым давлением, верхним двигателем водного потока или присасывающим действием атмосферы.

Основной силой, вызывающей поступление и передвижение воды в растении, является процесс транспирации, в результате которого возникает градиент водного потенциала. Градиент водного потенциала между клеткой и окружающим пространством создает движущую силу потока воды через мембрану. Если окружающая клетку среда представляет собой гипертонические, более концентрированные, чем клеточный сок, растворы, то вода станет выходить из клетки наружу. Это приведет к потере тургора клеткой, отделению плазмалеммы от клеточной стенки и обособлению протопласта — явлению плазмолиза.
Механизм, обеспечивающий поднятие воды по растению за счет корневого давления, — носит название нижнего концевого двигателя водного тока.
Корневое давление создается при переходе воды из коры корня в сосудистую систему корня при прохождении воды через пропускные клетки перицикла, из которых вода под давлением как бы впрыскивается в сосуды ксилемы. Доказательством этого служат явления гуттации и «плача растений».
Вода, поглощенная корневыми волосками и другими клетками эпидермиса, из клеток внешней части корня перемещается к ксилеме, занимающей центральную часть корня. Главным путем диффузии воды во внешней части корня служит аполаст - непрерывная совокупность клеточных стенок. Однако в эндодерме (цилиндрическом слое клеток, окружающем проводящую ткань) свободная диффузия по клеточным стенкам наталкивается на преграду - водонепроницаемый пробковый слой пояска Каспари. Вода должна изменить здесь свой путь и пройти сквозь мембрану и протопласт клеток эндодермы, играющей, таким образом, роль осмотического барьера между корой корня и его центральным цилиндром. У однодольных пробковеют также и внутренние тангенциальные стенки клеток, но эти стенки пронизаны порами, по которым, как по каналам, может проходить вода.
По ксилеме вода поднимается в надземные части растения. Ксилема состоит из нескольких типов клеток. Вода движется в ней главным образом по сосудам и трахеидам. И те и другие клетки прекрасно приспособлены для этой цели: они вытянуты в длину, лишены живого содержимого и внутри полые, т. е. это как бы трубки для воды. Одревесневшие вторичные клеточные стенки достаточно прочны на разрыв, чтобы выдерживать огромную разность давлений, возникающую при подъеме воды к вершинам высоких деревьев. Торцевые, а иногда и боковые стенки члеников сосудов перфорированы; сосуды, состоящие из соединенных конец в конец члеников, образуют длинные трубки, по которым легко проходит вода с растворенными в ней минеральными веществами. В трахеидах нет перфораций, и вода, для того чтобы попасть из одной тра-хеиды в другую, должна пройти через их торцевые стенки; однако трахеиды - очень длинные клетки, а потому и эта конструкция достаточно хорошо приспособлена для проведения воды.
У цветковых растений есть и сосуды, и трахеиды; у более примитивных форм сосудов, как правило, нет.
Листовые жилки, состоящие из тяжей ксилемы и флоэмы, образуют в листе настолько густую сеть, что любая его клетка оказывается достаточно близко от источника воды. Из ксилемы вода диффундирует в стенки клеток мезофилла. Таким образом, вода в жидкой фазе заполняет весь путь от почвы - через корень и стебель - до клеток мезофилла в листе.
Новое в блогах
Суммарный поток воды направлен всегда в сторону меньшего водного потенциала, т. е. максимален в почве, несколько ниже в клетках корня и самый низкий в клетках, примыкающих к эпидермису листа. Малая величина показателя в этих последних клетках объясняется главным образом испарением воды с поверхности листа.
Вода переходит из растения в окружающий воздух главным образом в парообразном состоянии. В мезофилле листа имеются обширные межклеточные пространства, и каждая клетка мезофилла хотя бы одной своей стороной граничит с таким межклетником. Вследствие испарения воды с влажных клеточных стенок воздух в межклетниках насыщен водяными парами, и часть этих паров теряется - выходит наружу. Поскольку у большинства растений клетки эпидермиса покрыты воскообразной водонепроницаемой кутикулой, водяные пары выходят из листа в атмосферу главным образом через устьица.
Гуттация — это выделение капельно-жидкой влаги листьями через гидатоды в условиях затрудненного испарения. Плач растения — это вытекание пасоки (воды с растворенными в ней минеральными веществами, находящейся в ксилеме) из стеблей растений со срезанными побегами. Гуттация обычно имеет место в условиях высокой влажности воздуха, когда «выключен»верхний концевой двигатель водного токатранспирация — физиологический процесс испарения воды надземными органами растений. Он осуществляется в основном из листьев через устьица и кутикулу.
Вода составляет до 95% массы растений, в ней или с ее использованием протекают все процессы жизнедеятельности. Поэтому вода необходимое условие для жизни организма. При недостатке воды у растения нарушается обмен веществ.
- Вода обеспечивает поток питательных и минеральных веществ по проводящей системе растения.
- Прорастание семян зависит от наличия воды.
- Вода участвует в процессе фотосинтеза.
- Водные растворы, наполняющие клетки и межклетники, обеспечивают растению упругость, таким образом растение сохраняет свою форму.
Растение обязательно должно поглощать воду. Иначе, рано или поздно, жизнь его прервется. Обычно растение поглощает воду исключительно своей корневой системой из почвы. В этом участвуют корневые волоски корней. Листья же через устьица испаряют воду. Смысл поглощения излишек воды, чтобы потом ее испарить, по большей части сводится к тому, что ток воды обеспечивает перенос веществ.
Если испарение воды растением превышает поступление воды, то у растения наблюдается увядание. Так нередко бывает днем, когда жарко. Ночью растение восполняет недостаток, так как испарение в это время суток снижено.
Вода в растение поглощается путем осмоса. При осмосе вода, в которой меньше растворенных веществ как бы засасывается в более насыщенные веществами растворы.
Как происходит водный обмен у растений: процессы и движение воды по растениях
Клеточные растворы растений более насыщенные, поэтому клетки впитывают воду.
В результате постоянного поглощения и испарения воды в растении существует постоянный водный обмен, включающий три этапа: поглощение воды корнями, передвижение ее по сосудам проводящей ткани, испарение воды листьями. Ток воды идет через все органы растения. Сколько растение всасывает воды, приблизительно столько оно его испаряет. Лишь доли процента от поступившей воды идут на синтез веществ. Это достаточно большие объемы воды. Так, например, только одно растение пшеницы в поле испаряет около 50 г воды в сутки.
Когда корни поглощают воду, они вместе с ней поглощают и растворенные минеральные соли. Когда вода испаряется, то соли в ней уже отсутствуют, они остаются в растении и используются в обмене веществ.
Водный ток идет снизу вверх. Его сила зависит от интенсивности всасывания корней и испарения листьями. Водный ток объединяет все органы растения, переносит различные соединения, питает клетки водой.
Новое в блогах
Нижний и верхний концевые двигатели. Корневое давление, его механизм и значение в жизни растений
В результате активной работы ионных на-сосов в корне и осмотического (пассивного) поступления во-ды в сосуды ксилемы в сосудах развивается гидростатическое давление, получившее название корневого давления. Оно обеспе-чивает поднятие ксилемного раствора по сосудам ксилемы из корня в надземные части. Показано, что у растений, обитаю-щих в холодных и плохо аэрируемых почвах, а также в резуль-тате действия ядов и ингибиторов корневое давление снижено. Механизм поднятия воды по растению вследствие разви-вающегося корневого давления называют нижним концевым двигателем.
Верхний концевой двигатель, обеспечиваю-щий передвижение воды вверх по растению, создается и под-держивается высокой сосущей силой транспирирующих клеток листовой паренхимы.
Транспирация - это физиологический процесс испарения во-ды растением. Основным органом транспирации является лист.
Растение имеет очень большую листовую поверхность, что создает огромную поверхность испарения. В результате потери воды клетками листьев в них снижается водный потенциал, т. е. возрастает сосущая сила. Это приводит к усилению поглощения клетками листа воды из ксилемы жилок и передвижению воды по ксилеме из корней в листья.
Сила верхнего концевого двига-теля будет тем больше, чем активнее транспирация. Верхний концевой двигатель может работать при полном отключении нижнего концевого двигателя, причем для его работы исполь-зуется не метаболическая энергия, а энергия внешней среды - температура и движение воздуха.
Предыдущая12345678910111213Следующая
ПОСМОТРЕТЬ ЕЩЕ:
Клетки растений используют осмос для увелечения обьема вакуоли, чтобы она распирала стенки клетки(тугурное давление).Клетки растений делают это путем запасания сахарозы. Увеличивая или уменьшая концентрацию сахарозы в цитоплазме, клетки могут регулировать осмос. За счёт этого повышается упругость растения в целом. С изменениями тургорного давления связаны многие движения растений (например, движения усов гороха и других лазающих растений). Пресноводные простейшие также имеют вакуоль, но задача вакуолей простейших заключается лишь в откачивании лишней воды из цитоплазмы для поддержания постоянной концентрации растворённых в ней веществ.


Осмос через полупроницаемую мембрану. Частицы растворителя (синие) способны пересекать мембрану, частицы растворённого вещества (красные) - нет.
В этой главе мы разобрались с осмосом- одним из важнейших устройств для транспорта воды.
2.3. Транспирация, как механизм транспорта воды
Начало изучении транспирации началось с XVIII века, но научный подход к обьянению этого явления начался в середине XIX века.Одним из первым исследователям транспирации был Г.Моль (1856), который докозал, что величина устьичных отверстий определяется тургором замыкающих клеток и зависит от света, тепла и влажности воздуха. Также он проводил эксперементы в результате которых он доказал, что рисутствие в замыкающих клетках хлоро-пластов, синтезирующих осмотические вещества, и таким образом влияющих на работу устьиц и на транспирацию.
Исследовательская работа:движение воды в растениях
Другими учеными, изучающими транспирацию были. Унгер (1857), опубликовавший в 1862 г. большую работу о транспирации. С. Швенденер (1883) высказал мысль, что устьица обеспечивают не только испарение, но и усвоение СОг- Представление об активной роли замыкающих, а не прилегающих к ним эпидермальных клеток, как это считал Дейтгеб (1886), окончательно утвердил сын Чарлза Дарвина Ф. Дарвин (1898). Действие различных лучей спектра на работу устьиц первым исследовал Коль (1895). Он установил, что красные и синие лучи, т. е. лучи, поглощенные хлорофиллом, вызывают открывание устьиц. Кроме устьичной транспирации в 1878 г. была обнаружена еще и кутикулярная (Хенель). Определения количеств испаряемой воды (Га-берландт, 1877; Хенель, 1879, 1880) показали, что эта величина различна в зависимости от природы самого растения и условий его произрастания. Но обо всем по порядку.
Транспирация — процесс потери растениями воды в виде пара. Основной орган для транспирации — лист. Аналогом транспирации является физический процесс испарения. Транспирация- один из важнейших факторов водного режима растений, так как испарение создает определенный энергетический импульс, который является причиной передвижения воды по растению. В связи с этим транспирация определяет скорость поглощения воды растением и вызывает водный дефицит в листьях. Также из-за потери воды в ходе процесса транспирации возрастает сосущая сила. Это приводит к усилению поглощения клетками листа воды из сосудов и передвижению воды по ксилеме из корней в листья. Процесс транспирации может осуществляться лишь при полном отключении нижнего концевого двигателя для работы верхнего концевого двигателя.Науке известно два вида транспирации: кутикулярная и устьичная. Кутикулярная транспирация представляет собой механтзм, в котором листья должны иметь однослойный эпедермис, внешние стенки клеток которого покрыты кутикулой и воском, образующие барьер на пути движения воды. На поверхности листьев часто развиты волоски, которые также влияют на водный режим листа, так как снижают скорость движения воздуха над его поверхностью и рассеивают свет, тем самым уменьшают потери воды за счет транспирации.Устьичная транспирация. Транспирация через устьица идет почти с такой же скоростью, как и обычная.Транспирация спасает растения от перегрева.
Итак, подведем итог. Мы узнали что транспирация в своей основе является физическим процессом испарения, который контролируется физическими факторами. Однако транспирация — это и физиологический процесс, который находится под влиянием таких внутренних факторов, как строение и расположение листьев, поведение устьиц. Транспирация обычно происходит в две стадии: испарение воды из клеточных стенок в межклетники и диффузия водяного пара в наружную атмосферу

2.4. Корневое давление, как механизм транспорта воды
Корневое давление – один из наиболее изучаемых физиологических процессов. Впервые оно было описано английским ученым С. Гельсом в 1727 году. Интерес к изучению механизмов этого явления не ослабевает. В России одним из первых подробно исследовал экссудацию знаменитый физиолог Д. А. Сабинин. Вначале он рассматривал корневую систему как осмотический аппарат, но позднее пришел к выводу о том, что экссудация является сложным физиологическим процессом, тесно связанным с метаболизмом клеток и их полярностью. В настоящее время все большее число ученых- физиологов растений признает осмотическую концепцию экссудации недостаточной для объяснения работы корня. на основании проведенных многочисленных опытов показано, что деятельность корневой системы складывается из двух составляющих: осмотической и метаболической. Корневое давление – это давление в проводящих сосудах корней, обеспечивающее наряду с транспирацией снабжение водой надземных органов. Оно возникает главным образом в результате повышения осмотического давления в сосудах корня (обычно 1-3 атмосферы) над осмотическим давлением почвенного раствора как следствие активного выделения клетками корня минеральных и органических веществ в сосуды. Обратному току жидкости из сосудов препятствует слой клеток эндодермы с опробковевшими (пропитанными суберином) оболочками. Результатом высокого корневого давления является «плач» растений, а также хорошо известное жителям Беларуси, США, Канады и других стран весеннее вытекание сока при надрезе стволов у березы и клена. Корневое давление имеет огромное значение также в поглощении воды растением в весеннее время до распускания листьев и у проростков при подземном прорастании. Велика его роль в восстановлении разорванных тяжей в сосудах ксилемы, по которым идет восходящий ток воды (от корней – к листьям). В ночные часы корневое давление ликвидирует возникший за день водный дефицит. Присасывающее действие транспирации передается корням в форме гидродинамического натяжения, которое связывает между собой работу этих двух двигателей.

Опыт, демонстрирующий корневое давление
3. Заключение
ДОБАВИТЬ КОММЕНТАРИЙ
[можно без регистрации]
перед публикацией все комментарии рассматриваются модератором сайта — спам опубликован не будет