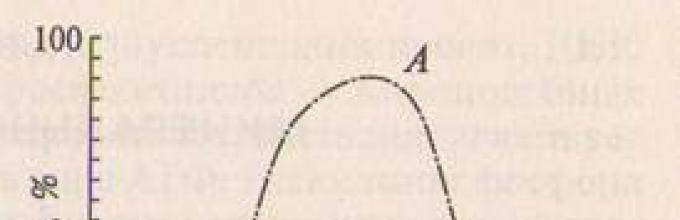
В надземные части растения вода поднимается по ксилеме.
У хвойных она движется по трахеидам, у лиственных - по сосу-
дам и трахеидам. Эти клетки хорошо приспособлены для этой цели: они вытянуты в длину, лишены цитоплазмы и внутри полые, т.е. это как бы трубки для воды. Одревесневшие вторичные клеточные стенки достаточно прочны на разрыв, чтобы выдерживать огром-ную разность давлений, возникающую при подъеме воды к вер-шинам высоких деревьев. В ксилеме взрослых деревьев воду прово-дят в основном ее периферические слои - заболонь.
Движущей силой восходящего тока воды в проводящих эле-ментах ксилемы является градиент водного потенциала через рас-тение от почвы до атмосферы. Он поддерживается градиентом ос-мотического потенциала в клетках корня и транспирацией. Для поглощения воды корнями требуются затраты метаболической энергии. На транспирацию используется энергия солнечной ради-
ации. Транспирация - главная движущая сила восходящего тока воды, так как благодаря ей в ксилеме возникает отрицательное давление, т.е. натяжение. В силу сцепления (когезии) молекул воды друг с другом и действия сил прилипания (адгезии) ее к гидро-фильным стенкам сосудов столб воды в ксилеме непрерывен. Со-четание транспирации, когезии и натяжения обусловливает подъем воды в стволах высоких деревьев. У большинства древесных расте-ний водный ток в стволе передвигается по спирали. Это связано с макростроением древесного ствола. Линейная скоростьвосходящ-его тока колеблется от 1 - 6 м/ч у хвойных и рассеянно-сосуди-тых пород до 25 - 60 м/ч у кольцесосудистых. Он обеспечивает все живые клетки растения водой и минеральными элементами.
Содержание воды в древесине у большинства древесных растен-ий увеличивается от внутренней части ствола к внешней и от основания ствола к его вершине. В пределах кроны количество воды увеличивается от вершины к основанию. Резкие изменения овод-ненности древесины наблюдаются в течение года. Так, у хвойных древесных растений самая низкая влажность отмечается в летние месяцы, а самая высокая - зимой. Влажность ядровой древесины практически не меняется и остается самой низкой. У лиственных древесных пород отмечены два периода пониженной влажности- летний и во второй половине зимы и два повышенной - весен-ний во время сокодвижения и зимний - в первой половине зимы. В течение суток летом наиболее высокая влажность отмечается утром, а низкая - в полдень.
10.4. Транспирация
Основной орган транспирации - лист. В результате потери воды клетками листьев в них снижается водный потенциал, т.е. возрас-тает сосущая сила. Таким образом, верхний концевой двигатель, обеспечивающий передвижение воды вверх по растению, созда-ется и поддерживается высокой сосущей силой транспирирую-щих клеток листовой паренхимы. Физиологическая роль транспи-рации сводится к следующему: 1) повышает сосущую силу испа-ряющих клеток и создает непрерывный водный ток по растению;
2) способствует передвижению воды и растворенных в ней мине-ральных и частично органических веществ от корней к надзем-ным частям растения; 3) защищает листья от перегрева прямыми солнечными лучами; 4) препятствует полному насыщению кле-ток водой, так как при небольшом водном дефиците (до 5%) оптимизируется целый ряд процессов метаболизма.
Транспирация бывает устьичная, кутикулярная и корковая (пе-ридермальная). Испарение воды как физическое явление, т.е. пе-реход воды из жидкого состояния в парообразное, происходит в межклетниках листа с поверхности клеток мезофилла. Образую-щийся пар через устьица выделяется в атмосферу. Это устьичная транспирация.
Устьица - основные проводящие пути для водяного пара, СО, и О,. Они могут находиться на обеих сторонах листа, но есть виды, у которых устьица располагаются только на нижней стороне лис-та. В среднем число устьиц колеблется от 50 до 500 на 1 мм". Тран-спирация с поверхности листа через устьица идет почти с такой же скоростью, как с поверхности чистой воды.
Потери водяного пара через кутикулу листа при открытых ус-тьицах обычно очень незначительны по сравнению с общей транс-пирацией. Но если устьица закрыты, например во время засухи, кутикулярная транспирация приобретает важное значение в вод-ном режиме многих растений. Кутикулярная транспирация зави-
сит от толщины слоя кутикулы и сильно варьирует у разных видов.
У молодых листьев она составляет около половины от общей транс-пирации, у зрелых листьев, с более мощной кутикулой, - не превышает 10 %.
Некоторое количество воды выделяется в результате транспи-рации почек и репродуктивных органов. Иногда эти потери могут быть значительными: например, корзинки подсолнечника, коро-бочки мака и плоды перца транспирируют сильнее, чем листья данных растений в тех же условиях. Вода испаряется с поверхнос-ти ветвей и стволов древесных растений через чечевички и окру-жающие их слои пробки. Это пробковая, или перидермальная, транс-пираиия. Вследствие транспирации ветвей и почек в зимнее время нередко наблюдаются случаи, когда значительные потери воды приводят к суховершинности древесных растений.
Скорость транспирации и газообмен в целом регулируются ус-тьицами. Степень раскрытости устьиц зависит от освещенности, оводненности тканей листа, концентрации СО2 в межклетниках и " .других факторов. В зависимости от факторов, запускающих двига-тельный механизм (свет или начинающийся водный дефицит в тканях листа), различают фото- и гидроактивное движение усть-иц. На свету в хлоропластах замыкающих клеток начинается фото-синтез, что вызывает уменьшение содержания СО2, накопивше-гося за ночь в клетках. При этом происходит накопление АТФ и, превращение крахмала в сахар,за счет которого активизируются
ионные насосы, перекачивающие калий из соседних клеток. Бла-годаря этому резко повышается сосущая сила устьичных клеток, всасывающих воду и увеличивающих тургор. Все это способствует открыванию устьиц. При возникновении водного дефицита повы-шается содержание одного из гормонов - абсцизовой кислоты; , под ее действием происходит отток и других растворенных ве-ществ, что приводит к закрыванию устьиц. Этот механизм позво-ляет защитить растение от излишней потери воды.
Показателем транспирации является ее интенсивность - коли-честно воды, испаренной в единицу времени на единицу сырой или сухой массы или поверхности листа (мг/дм2ч, г/м2ч или мг/г ч).
Количество граммов сухой массы, образовавшейся при испаре-нии 1 л воды, называется продуктивностью транспирации. Под эвапотранспирацией понимается сумма транспирационных потерь всеми растениями сообщества плюс физическое испарение (эва-порация) с поверхности почвы и растений, в частности со ство-лов и ветвей деревьев. Для лесных площадей центральных районов европейской части России транспирация древостоя в среднем соста-вляет 50 - 60 % эвапотранспирации, напочвенного покрова - 15- " 25%, испарение с поверхности почвы и растений - 25 - 35%.
Транспирацию кроны дерева используют для подсушки древе-сины после рубки. Свежесрубленная древесина ряда древесных пород (лиственница, береза, осина и др.) настолько тяжела, что тонет при сплаве. Вместе с тем более сухая и, следовательно, бо-лее легкая древесина тех же пород с успехом сплавляется на дале-кие расстояния. Для подсушки срубленное дерево оставляют ле-жать в лесу с кроной в течение 10 - 15 суток. Дерево продолжает жить за счет внутренних запасов воды и питательных веществ, а листья транспирировать. Количество свободной воды в стволе по-нижается. Уменьшение массы 1 мз древесины за указанное время составляет 25 - 30%, что резко повышает ее плавучесть. Облегча-ются также ее трелевка и транспортировка. Известно, что после сплава предварительно подсушенная древесина быстрее сохнет, чем не подсушенная до сплава.
На интенсивность транспирации влияет ряд факторов: доступ-ность воды корням растений, влажность воздуха, температура, ветер. При недостатке воды в почве интенсивность транспирации древесных растений заметно снижается. На затопленной почве этот процесс, несмотря на обилие воды, также снижен у деревьев при-мерно в 1,5 - 2 раза, что связано с плохой аэрацией корневых систем. Уменьшается транспирация и при сильном охлаждении почвы в связи со снижением скорости поглощения воды. Недоста-ток или избыток воды, засоление или холодная почва действуют на интенсивность транспирации не сами по себе, а через их вли-яние на поглощение воды корневыми системами.
Свет и влажность воздуха сильно воздействуют на транспира-цию. Свет увеличивает открытость устьиц. Интенсивность транс-пирации даже на рассеянном свету повышается на 30 - 40%, В темноте растения транспирируют в десятки раз слабее, чем при полном солнечном освещении. Повышение относительной влаж-ности приводит к резкому снижению интенсивности транспира-ции всех пород. Согласно закону Дальтона, количество испаряе-мой воды прямо пропорционально дефициту насыщения воздуха водяными парами.
Температура воздуха действует на транспирацию прямо и кос-венно. Прямое воздействие связано с нагреванием листа, а кос-венное - через изменение упругости паров, насыщающих про-странство. При повышении температуры количество пара в возду-хе уменьшается и транспирация усиливается. Ветер способствует повышению транспирации благодаря уносу паров воды от листь-ев, создавая недонасыщение воздуха у их поверхности.
В природе всегда действует комплекс факторов. В ходе дня ме-няются освещенность, температура и влажность воздуха, что ве-дет к изменению интенсивности транспирации (рис. 10.2). При умеренных температуре и влажности содержание воды в листьях уменьшается незначительно - на 10 - 15%. В жаркий день овод-ненность листьев снижается по сравнению с нормой до 25% и более.

Рис. 10.2. Дневной ход транспира-ции при различной влагообеспе-ченности растений:
А - испарение со свободной водной по-верхности; Б - транспирация при дос-таточной влагообеспеченности; В - при недостатке влаги в полдень; Г - при глубоком водном дефиците; Д - во вре-мя длительной засухи.
Различают дневной и остаточный водный дефицит. Дневной водный дефицит наблюдается в полуденные часы летнего дня. Как правило, он существенно не нарушает жизнедеятельность расте-ний. Остаточный водный дефицит наблюдается на рассвете и сви-детельствует о том, что водные запасы листа за ночь восстанови-лись лишь частично вследствие низкой влажности почвы. При этом растения сначала сильно завядают, а затем при длительной засухе могут погибнуть.
1. Из чего складывается водный режим растения?
2. Как происходит поглощение воды корнями?
3. В чем проявляется корневое давление?
4. Какие формы почвенной влаги доступны для растения?
5. Как осуществляется подъем воды к вершинам высоких деревьев?
6. Что такое транспирация и какая она бывает?
7. Как растение регулирует транспирацию?
8. Какие факторы внешней среды оказывают влияние на интенсив-ность транспирации?
МИНЕРАЛЬНОЕ ПИТАНИЕ.
Вода, поглощенная клетками корня, под влиянием разности водных потенциалов, которые возникают благодаря транспирации, а также силе корневого давления, передвигается до проводящих путей ксилемы.
Согласно современным представлениям, вода в корневой системе передвигается не только по живым клеткам. Еще в 1932 г. немецкий физиолог Мюнх развил представление о существовании в корневой системе двух относительно не зависимых друг от друга объемов, по которым передвигается вода, - апопласта и симпласта.
Апопласт - это свободное пространство корня, в которое входят межклетные промежутки, оболочки клеток, а также сосуды ксилемы.
Симпласт - это совокупность протопластов всех клеток, отграниченных полупроницаемой мембраной. Благодаря многочисленным плазмодесмам, соединяющим между собой протопласт отдельных клеток, симпласт представляет единую систему.
Апопласт, по-видимому, не непрерывен, а разделен на два объема. Первая часть апопласта расположена в коре корня до клеток эндодермы, вторая - по другую сторону клеток эндодермы, и включает в себя сосуды ксилемы. Клетки эндодермы благодаря пояскам
Каспари представляют как бы барьер для передвижения воды по свободному пространству (межклетникам и клеточным оболочкам). Для того чтоб попасть в сосуды ксилемы, вода должна пройти через полупроницаемую мембрану и главным образом по апопласту и лишь частично по симпласту.
Однако в клетках эндодермы передвижение воды идет, по-видимому, по симпласту. Далее вода поступает в сосуды ксилемы. Затем передвижение воды идет по сосудистой системе корня, стебля и листа.
Из сосудов стебля вода движется через черешок или листовое влагалище в лист. В листовой пластинке водопроводящие сосуды расположены в жилках. Жилки, постепенно разветвляясь, становятся более мелкими. Чем гуще сеть жилок, тем меньшее сопротивление встречает вода при передвижении к клеткам мезофилла листа.
Иногда мелких ответвлений жилок листа так много, что они подводят воду почти к каждой клетке. Вся вода в клетке находится в равновесном состоянии.
Иначе говоря, в смысле насыщенности водой, имеется равновесие между вакуолью, цитоплазмой и клеточной оболочкой, их водные потенциалы равны. Вода передвигается от клетки к клетке благодаря градиенту сосущей силы.
Вся вода в растении представляет единую взаимосвязанную систему. Поскольку между молекулами воды имеются силы сцепления (когезия), вода поднимается на высоту значительно большую 10 м. сила сцепления увеличивается, так как молекулы воды обладают большим сродством друг к другу. Силы сцепления обладают и между водой и стенками сосудов.
Степень натяжения водных нитей в сосудах зависит от соотношения процессов поглощения и испарения воды. Все это позволяет растительному организму поддерживать единую водную систему и не обязательно восполнять каждую каплю испаряемой воды.
В том случае, если в отдельные членики сосудов попадает воздух, они, по-видимому, выключается из общего тока проведения воды. Таков путь передвижения воды по растению (рис. 2).
Скорость перемещения воды по растению в течение суток изменяется. В дневные часы она на много больше. При этом разные виды растений различаются по скорости передвижения воды. Изменение температуры, введение метаболических ингибиторов не влияют на передвижение воды.
Вместе с тем этот процесс, как и следовало ожидать, очень сильно зависит от скорости транспирации и от диаметра водопроводящих сосудов. В более широких сосудах вода встречает меньшее сопротивление. Однако надо учитывать, что в более широкие сосуды могут попасть пузырьки воздуха или произойти какие-либо иные нарушения тока воды.
Рис.2. Путь воды в растении.
Включает в себя такие понятия, как поступление, движение воды в растениях и испарение ее.
Вода необходима растениям
Передвигается вода в растениях по клеткам коровой паренхимы до центрального цилиндра корня, затем по проводящей системе до листовой паренхимы и, наконец, по клеткам листовой паренхимы. На первом участке пути вода передвигается благодаря повышению сосущей силы клеток корня.
 Движение воды от корневого волоска в сторону центральных сосудов
Движение воды от корневого волоска в сторону центральных сосудов
Этот отрезок пути очень небольшой (доли миллиметра), но передвижение воды по этому участку очень затруднено, так как воде приходится преодолевать сопротивление слоев живой протоплазмы . Это сопротивление примерно равно 1 атм на 1 мм пути, поэтому передвижение воды по живым клеткам на более значительные расстояния не обеспечивало бы потребности растения в воде.
Действительно, растения , у которых не развита проводящая система , например мхи, (подробнее: ) имеют незначительные размеры и приспособлены к жизни только во влажных условиях. У наземных растений в процессе эволюции образовалась проводящая ткань, которая устанавливает сообщение между всасывающими воду корнями и испаряющими воду листьями.
Проводящая воду ткань
Проводящая воду ткань состоит из сосудов, или трахей , и трахеидов ; она начинается в центральном цилиндре корня, проходит через весь корень и стебель и заканчивается в виде тончайших разветвлений - жилок, пронизывающих всю листовую паренхиму.
Сосуды представляют собой мертвые трубки, образовавшиеся из живых клеток. В сосудах сохраняются поперечные перегородки на разном расстоянии (от нескольких миллиметров до метра в зависимости от вида растения) одна от другой.
Исчезновение перегородок даже на небольшом расстоянии в тысячи раз ускоряет передвижение воды. Трахеиды это длинные мертвые клетки с заостренными концами. При образовании сосудов и трахеид происходит утолщение и одревеснение их оболочек, вследствие чего они не сдавливаются под давлением окружающих их живых паренхимных клеток.
 Движение воды в древесных растениях
Движение воды в древесных растениях
Одревеснение, однако, никогда не бывает сплошным: на стенке сосудов остаются тонкие места - поры, по которым вода может перемещаться не только вверх по сосудам, но и в радиальном направлении.
Подъем воды по сосудам
Подъем воды по сосудам можно доказать следующим опытом. Если у срезанной и поставленной в воду ветки снять кольцо коры выше уровня воды, листья ее не завянут, так как сосуды расположены в древесине.
Движение воды по сосудам чаще всего направлено снизу вверх и называется поэтому восходящим током .

Последний отрезок пути водного тока по листовой паренхиме идет по живым
клеткам
. Вода передвигается осмотическим путем по клеткам мезофилла листа до последних клеток, граничащих с межклеточниками
. Этот отрезок пути, так же как и первый, очень короткий.
Если срезанную ветку растения герметически закрепить в стеклянной трубке, заполненной водой, и нижний конец ее опустить в сосуд со ртутью, то при испарении веткой воды ртуть в трубке будет подниматься.
Из этого опыта ясно, что передвижение воды по растению обусловлено главным образом транспирацией , (подробнее: ), а не только корневым давлением.
При испарении воды с поверхности листьев в клетках возникает сосущая сила. Величина ее тем больше, чем меньше воды остается в клетках листа. Эта возникающая сосущая сила поддерживает постоянное передвижение воды в растении.
Силы, приводящие воду в движение
Таким образом, силы, приводящие воду в движение , находятся по концам проводящей системы: нагнетающий воду корень, работа которого получила название нижнего концевого двигателя , и сила присасывания воды листьями - верхний концевой двигатель .
Оба двигателя действуют в одном направлении и могут заменять и дополнять друг друга. Во время сильной инсоляции летом и при, засухе водоснабжение растения идет за счет присасывающего действия транспирации.
Корневое давление
Когда же почва богата водой, а воздух водяными парами, подъем воды обеспечивается силой корневого давления, (подробнее: ). Следовательно, в зависимости от условий внешней среды главная роль принадлежит то одному, то другому концевому двигателю.
Водные нити не рвутся под влиянием своей тяжести, несмотря на то, что при сильной они находятся в состоянии натяжения. Это объясняется силой сцепления молекул воды, достигающей 300-350 атм, а так как в сосудах нет воздуха, то целостность водного тока не прерывается.
Скорость водного тока
Скорость водного тока зависит от строения проводящих воду элементов. Вода быстрее передвигается по сосудам, причем скорость движения ее зависит от диаметра сосудов: чем он меньше, тем медленнее будет передвигаться вода.
Движение воды в растениях происходит благодаря работе двух концевых двигателей, верхнего и нижнего, и сил сцепления, обеспечивающих целостность водных нитей.
МИНИСТЕРСТВО ОБРАЗОВАНИЯ РЕСПУБЛИКИ БАШКОРТОСТАН
МУНИЦИПАЛЬНОЕ БЮДЖЕТНОЕ ОБРАЗОВАТЕЛЬНОЕ УЧРЕЖЕНИЕ
СРЕДНЯЯ ОБЩЕОБРАЗОВАТЕЛЬНАЯ ШКОЛА №85
ОРДЖОНИКИДЗЕВСКОГО РАЙОНА ГОРОДСКОГО ОКРУГА ГОРОД УФА
РЕСПУБЛИКИ БАШКОРТОСТАН
Секция: «Окружающий мир»
Тема: Движение воды в растениях
ученица 2 «Б» класса
Научный руководитель : Сафарова Л.М
учитель начальных классов
Уфа -2013
Оглавление
Введение…………………………………………………..с.3
Глава I. Основы поглощения воды……………………..с. 4-6
1.1 Корневая система как орган поглощения воды
1.2 Основные двигатели водного тока
Глава II . Транспирация………………………………….. с.7-9
2.1 Назначение транспирации
Глава III . Адаптация к дефициту воды…………………с.10-11
Заключение……………………………………………….с.12
Список литературы………………………………………с.13
Введение
Если посмотреть на фотографию нашей планеты, полученную из космоса, сразу видно обилие голубого цвета на земном шаре. Это - вода, занимающая три четверти поверхности Земли. Вода - все знают, как она проста. Но за этой кажущейся простотой - свойства самого удивительного и замечательного вещества на Земле.
Данная тема достаточно актуальна , так как за последние десятилетия возросла скорость изменений, относящихся к различным явлениям жизни всего мирового сообщества, в том числе и по отношению к воде.
Цель работы: изучить движение воды в растениях.
Постановка указанной цели определяет круг задач:
Сбор и анализ литературы по данному исследованию;
Провести анализ литературы;
Провести опытно – экспериментальную работу.
Объект исследования : вода
Предмет исследования : движение воды в растениях
Практическая ценность работы состоит в широких возможностях применения основных выводов настоящего исследования на уроках, классных часах, внеклассных мероприятиях.
Глава I. Основы поглощения воды
Живые растительные клетки на 80-90 процентов состоят из воды. Даже клетки сухих семян, в которых приостановлена жизнь, содержат 10 процентов воды. Листья растений постоянно испаряют воду, особенно днем. Это происходит потому, что их поверхность усеяна многочисленными микроскопическими отверстиями - устьицами. Причем на нижней поверхности листа, их значительно больше, чем на верхней. Днем устьица открыты и водяные пары выходят из листа. Ночью же они закрываются, и испарение воды практически прекращается. Но и ночью растение медленно теряет воду. Она уходит, минуя устьица, через тонкую кожицу листа.
Водный ток обеспечивает связь между отдельными органами растений. Питательные вещества передвигаются по растению в растворенном виде. Насыщенность водой обеспечивает прочность тканей и сохранение структуры травянистых растений. Рост клеток идет главным образом за счет накопления воды в определенных ее частях.
Таким образом, вода обеспечивает протекание процессов обмена. Для нормальной жизнедеятельности клетка должна быть насыщенна водой.
Основным источником влаги является вода, находящаяся в почве, и основным органом поглощения воды является корневая система. Роль этого органа, прежде всего, заключается в том, что благодаря огромной поверхности обеспечивается поступление воды в растения из возможно большого объема почвы.
Корневая система как орган поглощения воды
Водную проблему растение решает с помощью хорошо развитой водопроводящей системы, которая начинается в корнях, поглощающих влагу из почвы, продолжается в трубках, подающих ее ко всем частям растения, и заканчивается испарением из листьев в воздух. Кажется, все просто. Однако механизм передвижения воды на самом деле сложен и не до конца еще изучен.
Рост корня, его ветвление продолжается в течение всей жизни растительного организма, то есть практически он не ограничен. Определение размеров корневых систем требует специальных методов. Оказалось, что общая поверхность корней обычно превышает поверхность надземных органов в 104-150 раз. При выращивании одиночного растения ржи было установлено, что общая длинна его корней достигает 600 км при этом на них образуется 15 миллиардов корневых волосков. Эти данные говорят об огромной потенциальной способности к росту корневых систем. Однако эта способность не всегда проявляется. При росте растений с достаточно большой густотой размеры корневых систем заметно уменьшаются.
Корневые волоски проникают в самые мелкие трещины почвы и, обнаружив хоть немного влаги, поглощают ее.
Важное значение для развития корневых систем имеет кислород. Именно недостаток кислорода является причиной плохого развития корневых систем на заболоченных почвах. Растения, приспособленные к росту на плохо продуваемых почвах, имеют в корнях систему межклетников, которые вместе с межклетниками в стеблях и листьях составляют единую вентиляционную систему.
Основные двигатели водного тока
Поглощение воды корневой системой идет благодаря работе двух двигателей водного тока: верхнего двигателя, испарения (транспирации), и нижнего двигателя, или корневого двигателя. Основной силой, вызывающей поступление и передвижение воды в растении, является сила испарения, в результате которой возникает градиент приток водного потенциала. Водный потенциал – это мера энергии, которую использует вода для передвижения. Водный потенциал и сосущая сила одинаковы по значению, но противоположны по знаку. Чем меньше насыщенна водой данная системы, тем меньше ее водный потенциал.
Таким образом, верхний двигатель водного тока в растении – это присасывающая сила листьев, и его работа мало связана с жизнедеятельностью корневой системы. Действительно, опыты показали, что вода может поступать в побеги и через мертвую корневую систему, причем в этом случае поглощение воды даже ускоряется.
Глава II . Транспирация
Транспирация определяется как испарение воды в атмосферу с листьев и стеблей живых растений. Растения впитывают влагу, содержащуюся в почве, через корни, причем эта вода может брать начало глубоко под землей. Так, например, зерновые растения имеют корни длиной до 2.5 метров, а корни некоторых растений пустыни уходят в землю на глубину 20 метров. Вода, которую выкачивают растения из-под земли, доставляет питательные вещества к листьям растений. Эта подкачка регулируется испарением воды через небольшие поры, которые расположены с обратной стороны листьев. Растение испаряет воду, когда влажность окружающего воздуха ниже, чем влажность воздуха в порах; в противном случае растение поглощает водяной пар из воздуха. Транспирация отвечает приблизительно за 10% всей испаряющейся влаги.
Для того чтоб наглядно рассмотреть этот процесс мы провели опыт. Для опыта нам понадобились: белые цветы, пищевые красители, ёмкости для воды, нож, вода.
Согласно плана работы:
Наполнили емкости водой
Всем цветам срезали стебли под углом 45градусов в теплой воде, для большей наглядности эксперимента у одного цветка стебель разрезали на 2 части, не до конца.
Поместили по одному цветку в каждую емкость с красителем, цветок с расщепленным стеблем поместили в 2 емкости с красителями красного и бирюзового цвета.
В течение 24 часов наблюдали за цветами (См. Таблица 1).
Таблица 1
№ п/п
Прошедшее время
Описание наблюдений
1 час
Центр цветка проявил чуть заметную окраску, в основном бирюзовый цвет.
2 часа
Стали проявляться прожилки на других цветах
10 часов
С первого взгляда видно, что цветы приобрели разую окраску (лучше всего проявляется синий и бирюзовый цвет).
18 часов
Хорошо видны прожилки. На кончиках лепестков появляются ярко выраженные пятна.
24 часа
Цветок с расщепленным стеблем окрашен наполовину, бирюзовый цвет видно хорошо, а красный очень плохо
Объяснение опыта:
Вода поступает в растение из почвы через корневые волоски и молодые части корней и по сосудам разносится по всей его надземной части. С передвигающейся водой разносятся по всему растению поглощенные корнем минеральные вещества (в проделанном опыте это видно по окрашенным лепесткам). Цветы, которые мы используем в эксперименте, лишены корней. Тем не менее, растение не теряет возможность поглощать воду. Это возможно благодаря процессу транспирации - испарению воды растением. Основным органом транспирации является лист. В результате потери воды в ходе транспирации в клетках листьев возрастает сосущая сила. Транспирация спасает растение от перегрева. Кроме того, испарение участвует в создании непрерывного тока воды с растворенными минеральными и органическими соединениями из корневой системы к надземным органам растения.
2.1. Назначение транспирации
В обычно протекающих процессах транспирация не является необходимой. Так если выращивать растения в условиях высокой и низкой влажности воздуха, то, естественно, в первом случае транспирация будет идти сознательно меньшей интенсивностью. Однако рост растений будет одинаков или даже лучше там, где влажность воздуха выше, а транспирация меньше. Известно, что большая часть всей поглощенной энергии тратится на транспирацию, которая в определенном объеме полезна растительному организму.
1.Транспирация спасает растение от перегрева, который ему грозит на прямом солнечном свете. Это особенно важно в связи с тем, что перегрев, разрушая хлоропласты, резко снижает процесс фотосинтеза. Именно благодаря высокой транспирирующей способности многие растения хорошо переносят повышенную температуру.
2.Транспирация создает непрерывный ток воды из корневой системы к листьям, который связывает все органы растения в единое целое.
3. С транспирационным током передвигаются растворимые минеральные и частично органические питательные вещества, при этом, чем интенсивнее транспирация, тем быстрее идет процесс передвижения.
Глава III . Адаптация к дефициту воды
Вода является необходимым условием существования всех живых организмов на Земле. Значение воды в процессах жизнедеятельности определяется тем, что она является основной средой в клетке, где осуществляются процессы, служит важнейшим исходным, промежуточным или конечным продуктом биохимических реакций. Особая роль воды для наземных организмов (особенно растений) заключается в необходимости постоянного пополнения ее, из-за потерь при испарении. Поэтому вся эволюция наземных организмов шла в направлении приспособления к активному добыванию и экономному использованию влаги. Наконец, для многих видов растений, животных, грибов и микроорганизмов вода является непосредственной средой их обитания.
Увлажненность местообитания и, как следствие, водообеснечение наземных организмов зависят, прежде всего, от количества осадков, их распределения по временам года, наличия водоемов, уровня грунтовых вод, запасов почвенной влаги и т.д.
Экологические группы растений по отношению к влаге и их адаптации к водному режиму. Высшие наземные растения, ведущие прикрепленный образ жизни, в большей степени, чем животные, зависят от обеспеченности воздуха влагой.
Гигрофиты - растения избыточно увлажненных местообитаний с высокой влажностью воздуха и почвы. Наиболее типичные гигрофиты - травянистые растения влажных тропических лесов и нижних ярусов сырых лесов в разных климатических зонах (чистотел большой, недотрога обыкновенная, кислица обыкновенная и др.), прибрежные виды (калужница болотная, плакун-трава, рогоз, камыш, тростник), растения сырых и влажных лугов, болот (белокрыльник болотный, сабельник болотный, вахта трехлистная, осоки), некоторые культурные растения.
Ксерофиты - растения сухих местообитаний, способные переносить продолжительную засуху, оставаясь физиологически активными. Это растения пустынь, сухих степей, саванн, сухих субтропиков, песчаных дюн и сухих, сильно нагреваемых склонов. Структурные и физиологические особенности ксерофитов нацелены на преодоление постоянного или временного недостатка влаги в почве или воздухе. Решение данной проблемы осуществляется тремя способами:
Эффективным добыванием (всасыванием) воды
Экономным ее расходованием
Способностью переносить большие потери воды
К группе ксерофитов относятся - растения с сочными мясистыми листьями или стеблями, содержащими сильно развитую водоносную ткань. Различают листовые суккуленты (агавы, алоэ, молодило, очитки) и стеблевые, у которых листья редуцированы, а надземные части представлены мясистыми стеблями (кактусы, некоторые молочаи и др.).
Корневая система поверхностная, мало - развитая, рассчитана на поглощение воды из верхних слоев почвы, увлажненных редко выпадающими дождями. В засуху корни могут отмирать, но после дождей быстро (за 2-4 дня) отрастают новые.
Мезофиты - занимают промежуточное положение между гигрофитами и ксерофитами. Они распространены в умеренно влажных зонах с умеренно теплым режимом и достаточно хорошей обеспеченностью минеральным питанием. К мезофитам относятся растения лугов, травянистого покрова лесов, лиственные деревья и кустарники из областей умеренно влажного климата, а также большинство культурных растений и сорняки. Для мезофитов характерна высокая экологическая пластичность, позволяющая им адаптироваться к меняющимся условиям внешней среды.
Специфичные пути регуляции водообмена позволили растениям занять самые различные по экологическим условиям участки суши. Многообразие способов приспособления лежит, таким образом, в основе распространения растений на Земле, где дефицит влаги является одной из главных проблем экологической адаптации.
Заключение
Из всего выше перечисленного можно вывести общее заключение, что при дефиците влаги растения могут адаптироваться т.е образование корневой системы, которая достигает влажных зон почвы; ограничение расхода воды на транспирацию; запасание воды в тканях растений.
Так как вода является основной составной частью растительных организмов. Вода - это та среда, в которой протекает все процессы обмена веществ.
Водный ток обеспечивает связь между отдельными органами растений. Питательные вещества передвигаются по растению в растворенном виде. Насыщенность водой, обеспечивает прочность тканей, сохранение структуры травянистых растений.
Таким образом, вода обеспечивает протекание процессов обмена, связь организма со средой. Для нормальной жизнедеятельности клетка должна быть насыщенна водой.
Основным источником влаги является вода, находящаяся в почве, и основным органом поглощения воды является корневая система. Роль этого органа прежде всего заключается в том, что благодаря огромной поверхности обеспечивается поступление воды в растения из возможно большого объема почвы.
Большая детская энциклопедия Том 9 «Растения и животные» [электронный ресурс] – ООО Мастермедиа 2006 г. 1 электронный оптический диск (CD - ROM ).

Высшее растение расчленено на органы, которые выполняют разные функции, но имеют много общих свойств, в числе которых можно назвать необходимость питательных, веществ и воды для протекания физиологических процессов. Так как вода поглощается не всеми органами, а в основном корневой системой, возникает необходимость ее передвижения по растению. Этот процесс составляет так называемый восходящий ток. Следует отметить, что это название отражает не направление, а характер передвижения и его локализацию в растении. Он проходит главным образом по мертвым тканям стебля или черешка - сосудам или трахеям у покрытосеменных растений и трахеидам у голосеменных. Однако эта локализация не абсолютна: вода способна перемещаться и по другим анатомическим элементам, например по флоэмной системе.

Вода с растворенным и в ней минеральным и веществами поднимается по сосудам древесины.

Если принять во внимание всю протяженность пути восходящего тока, то его можно будет разбить на два не равных по длине участка.
1. Мертвые гистологические элементы в середине проводящего пути сосуды или трахеиды. Протяженность этого участка значительна, но вода проходит по нему сравнительно легко, так как двигается пассивно по мертвым элементам, не испытывая с их стороны значительного сопротивления.

2. Живые клетки корня и листа, находящиеся в начале и в конце пути передвижения. Этот путь пространственно короток, но преодолевается он с большим трудом, так как клеточные мембраны препятствуют передвижению воды.
Передвижение воды в восходящем токе имеет важное значение в жизни растения. Этот ток снабжает водой все органы и ткани, приводя их в состояние тургора. Восходящий ток воды захватывает минеральные ионы, поглощенные корнем, переносит их и тем самым облегчает распределение (но не поглощение!) по растению.

Для того чтобы вода двигалась по растению (и не просто двигалась, а поднималась вверх), необходимы затраты некоторого количества энергии, точки приложения которой располагаются по концам тока, вследствие чего они получили название концевых двигателей.
Нижний концевой двигатель, или корневое давление. Его роль проявляется в основном при активном поглощении - нагнетании воды. При участии сократительных белков он не только подает воду в корневую систему, но и проталкивает ее дальше в сосуды корня и вверх по стеблю. Нагнетание воды
Активный энергозависимый процесс, который сильнее всего выражен в коре корня. Сила, развиваемая концевым двигателем, невелика (около 0,15 МПа), она может обеспечить подъем воды на высоту не более одного метра, т. е. достаточна для травянистых растений и небольших кустарников.

Симпласт - это система взаимосвязанных протопластов растения. Протопласты соседних клеток соединяются между собой плазмодесмами - цитоплазматическими тяжами, проходящими через поры в клеточных стенках. Вода с любыми растворенными в ней веществами, попав в протопласт одной клетки, может двигаться дальше по симпласту, не пересекая никаких мембран. Это движение иногда облегчается благодаря упорядоченному току цитоплазмы.

Апопласт - это система соприкасающихся клеточных стенок, образующая непрерывную сеть по всему растению. До 50% такого целлюлозного каркаса представляет собой как бы « свободное пространство», которое может быть занято водой. При ее испарении в межклетники с поверхности клеток мезофилла в непрерывном апопластном слое воды возникает натяжение, и весь он по механизму объемного потока подтягивается к месту убывания благодаря когезии (« сцеплению») водных молекул. В апопласт вода поступает из ксилемы.

Верхний концевой двигатель, или присасывающая сила транспирации. При постоянном испарении воды в листьях растений разбивается сосущая сила (1 - 1,5 МПа), отсасывающая воду от ближайших клеток и передающаяся последующим клеткам, по которым передвигается вода, вплоть до сосудов. В сосудах нет цитоплазмы, поэтому нет и осмотического давления, и всасывание жидкости происходит при участии всей величины сосущей силы. Она позволяет поднять воду на несколько метров, действуя подобно гидравлическому насосу. Этой силы достаточно для обеспечения водой кустарников и сравнительно небольших деревьев.

Поднятие воды по стволу дерева
Концевые двигатели могут поднять воду на высоту до 10 м. Но многие древесные растения имеют гораздо большую протяженность ствола, и тогда оба концевых двигателя не могут обеспечить подъем воды. У таких растений на помощь приходят силы сцепления между молекулами воды, которые очень велики и могут достигать 30 - 35 МПа. Этой силы достаточно для того, чтобы поднять воду на 1 - 2 км, что значительно превышает высоту любого дерева.
Силы сцепления молекул воды действуют только при определенных условиях: водные струи в сосудах должны идти непрерывно, без пузырьков воздуха. Если же в них попадает воздух, что возможно при их поранении или перерезке, передвижение воды прерывается. Этим объясняется завядание побегов древесных растений с листьями и цветками (например сирени), когда их после срезания помещают в воду не сразу, а спустя некоторое время.